即日起,分析测试中心将不定期推送冷冻电镜国内外应用案例,供各团队谋划使用冷冻电镜设备开展科研创新提供参考!
题目:Helicase-mediated mechanism of SSU processome maturation and disassembly
链接: DOI: 10.1038/s41586-025-09688-3

真核核糖体小亚基 (SSU) 组装需要 SSU 突起组,这是一种含有 RNA 伴侣 U3 小核仁 RNA (snoRNA) 的核仁前体。由于缺乏中间体,SSU过程组成熟、重塑、分解和RNA质量控制的潜在分子机制以及状态之间的转变仍然未知。
2025年10月29日,Rockefeller大学Sebastian Klinge团队在Nature上发表题为Helicase-mediated mechanism of SSU processome maturation and disassembly的研究文章,揭示了两种解旋酶 Mtr4-外泌体和 Dhr1 如何被控制以实现准确和单向的核糖体生物发生。研究表明,冗余系留的RNA外泌体不可逆的核糖体前RNA降解如何耦合SSU过程组转化为40S前颗粒,在此期间Utp14可以探测进化的表面,最终定位和激活Dhr1以展开U3 snoRNA并启动核仁40S前释放。这项研究强调了大型动态 RNA-蛋白质复合物的范式,其中不可逆的 RNA 降解驱动组成变化并传达这些变化以控制酶活性,同时保持整体质量控制。
RNA外切体的“不可逆降解驱动”作用
Mtr4-外切体以3'→5'方向降解5'外部转录间隔区(5'ETS),触发18S rRNA的A1位点切割,同时驱动SSU加工体拆解。外切体通过多价锚定网络(UTP18的AIM基序、RRP6的“lasso”结构域、UTP14的ERM1/2基序)稳定结合SSU加工体,确保降解与复合物拆解同步。遗传学实验证实,该锚定网络具有冗余性,四锚定位点全失活才会导致细胞致死。
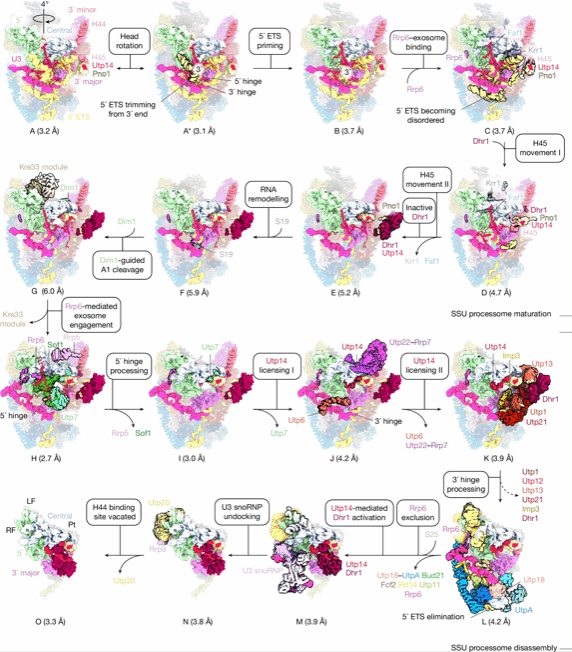
图1. SSU过程组成熟和拆卸路径
Dhr1解旋酶的“状态特异性激活”机制
UTP14作为核心调控因子,动态探测SSU加工体表面构象变化。早期态中,Dhr1受Pno1抑制元件与自身自抑制环调控处于失活状态。随着5'ETS降解与组装因子(如SOF1-UTP7、UTPB)逐步脱离,UTP14的B/D/E结构域释放,为Dhr1构建新结合位点并解除自抑制,使其解开U3 snRNA与前rRNA的碱基配对,促进核仁前40S颗粒释放。

图2. Utp14介导的Dhr1激活机制
核仁颗粒释放的“价态调控”基础
随着组装因子逐步脱离,SSU加工体多价相互作用减弱、“价态”降低,为其离开核仁进入后续成熟步骤创造条件。该研究建立了大型RNA-蛋白复合物动态调控的新范式:不可逆RNA降解不仅是“废物处理”,更是“分子开关”,驱动复合物组成变化,并通过UTP14等因子将变化信号传递给解旋酶,实现酶活性调控与质量控制的偶联。
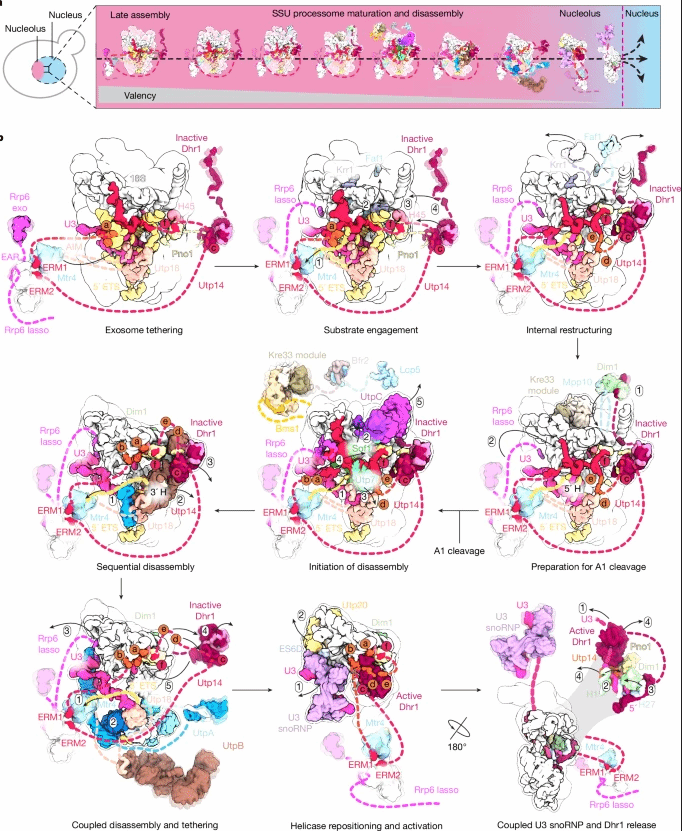
图3. RESC识别gRNA和mRNA过程中发生的状态变化
该研究建立了大型RNA-蛋白复合物动态调控的新范式:不可逆RNA降解不仅是“废物处理”,更作为“分子开关”驱动复合物组成变化,并通过Utp14等因子将变化信号传递给解旋酶,实现酶活性调控与质量控制的偶联。此外,结构数据还揭示了核仁颗粒释放的分子基础——随着组装因子脱离,SSU加工体的多价相互作用减弱,复合物“价态”降低,为其离开核仁进入后续成熟步骤创造条件。这些发现为理解核糖体组装障碍相关疾病的分子机制提供了重要基础。
本文来源 [ CNS分享微信公众号]
推文原链接:https://mp.weixin.qq.com/s/I4iZzIKM_62FIKRNpODufg
福建农林大学分析测试中心
2025年11月11日







