即日起,分析测试中心将不定期推送冷冻电镜国内外应用案例,供各团队谋划使用冷冻电镜设备开展科研创新提供参考!
题目: Evolutionary adaptations of doublet microtubules in trypanosomatid parasites
链接: DOI: 10.1126/science.adr5507

锥虫寄生虫是一类寄生性原生生物,包括 Leishmania、Crithidia 和 Trypanosoma 属。它们对人类健康具有重要影响,是利什曼病、锥虫病(非洲昏睡病)和恰加斯病的致病因子。这些寄生虫依赖鞭毛在宿主体内或昆虫媒介中存活。例如,Leishmania mexicana 的鞭毛型前鞭毛体(promastigote)在血食蠓(phlebotomine sand fly)体内生长,需要鞭毛运动才能从昆虫中肠迁移至口器,以便在叮咬哺乳动物宿主时完成传播。同样,Trypanosoma brucei 依靠鞭毛在采采蝇(tsetse fly)内运动,且在宿主体内若缺乏运动能力则会导致寄生虫死亡。
鞭毛的摆动由轴丝(axoneme)决定。轴丝是一个由双联微管(DMTs)组成的细胞内圆柱结构,沿鞭毛延伸。冷冻电子显微镜(Cryo-EM)最近已被用于解析多种单细胞生物的 DMT 结构,包括 Chlamydomonas reinhardtii(衣藻)【1,2】,Tetrahymena thermophila(四膜虫)【3】及各种哺乳动物纤毛细胞。这些研究阐明了装饰 DMT 内腔及外部的蛋白及其不同的周期性。然而,大部分结构蛋白的具体功能研究尚不完整,且往往在不同生物体中独立进行,难以直接比较。
为了解析锥虫家族 DMTs 的结构,我们使用了两种不感染人类的模型生物:双宿主的 L. tarentolae(感染爬行动物)和单宿主的 C. fasciculata(仅寄生于蚊科昆虫)。对于每个物种,我们通过机械力分离鞭毛轴丝的方法得到散开的 DMTs,进而运用单颗粒冷冻电镜(cryo-EM)技术解析 DMT 的 48 和 96 nm 重复单元的结构,分辨率高达 2.7 埃(Å)【图 1】。Alan Brown 和张锐课题组分别负责 Leishmania 和 Crithidia 的 DMT 的结构解析。Brown 实验室的博士后 Matthew Doran 和张锐实验室的博士生牛清薇为本文的共同第一作者。
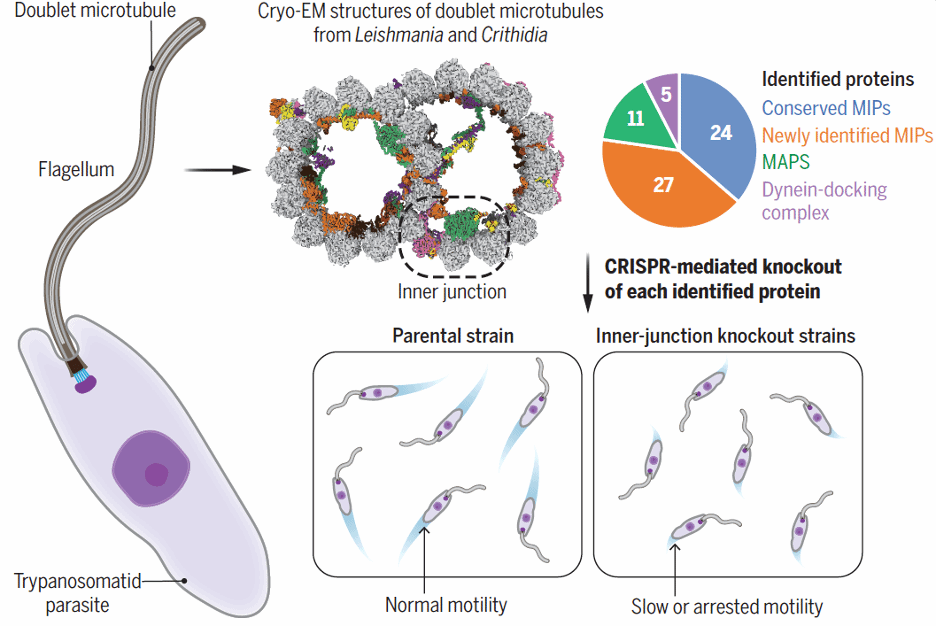
图 1:图示摘要: 锥虫双联微管的结构和功能分析
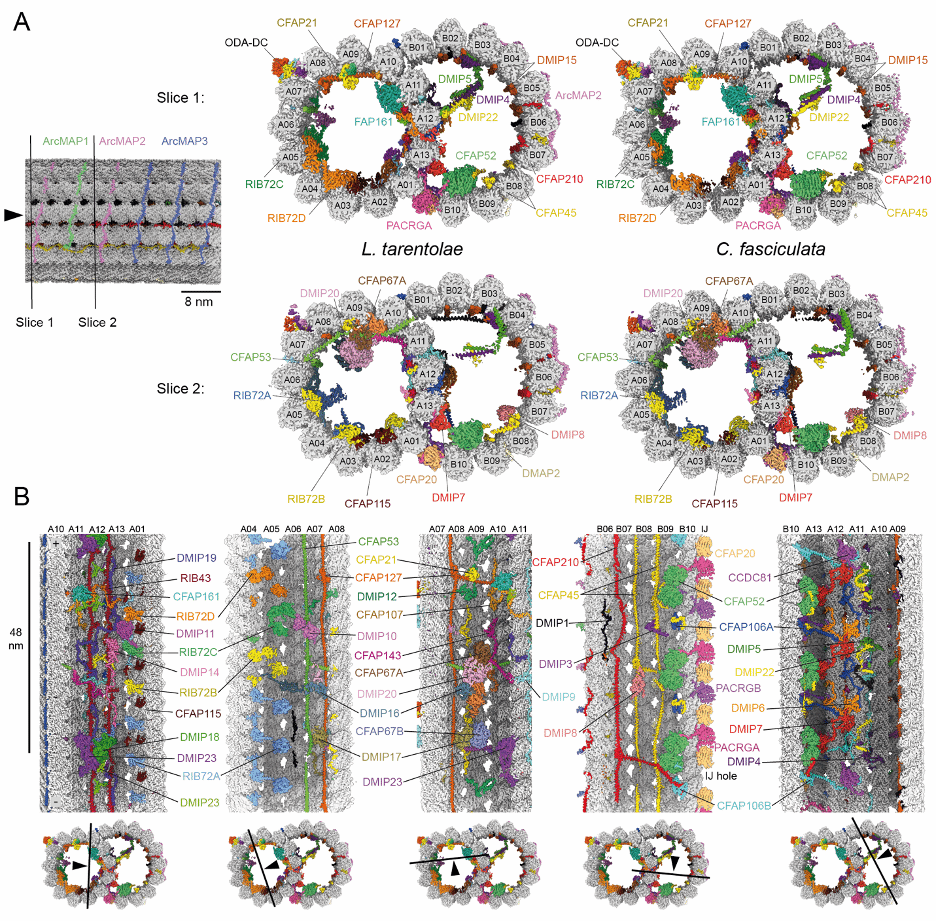
图 2:L. tarentolae 和 C. fasciculata 双联微管的 48 nm 重复单元的 cryoEM 结构
结果显示【图 2】,在单个 48 nm 的重复单元中,我们发现了 51 种不同的 MIPs,其中 27 种是锥虫寄生虫或更广泛的 kinetoplastids 特有的 MIPs,此外还发现了一种由五个亚基组成的 dynein-docking complex,以及一类新型的 MAPs,称为 ArcMAPs。
研究还发现,一些 MIPs 来自旁系同源基因(paralogous genes)【图 3】。这些同源蛋白在单个重复单元内的存在形成了一种新的周期性排列模式,使得 DMT 的 MIP 网络比以往解析的 DMT 结构更为复杂。此外,这些旁系同源蛋白还允许 DMT 形成近端-远端(proximal-distal)非对称性,例如,锥虫寄生虫的动力蛋白对接复合物含有特定的旁系同源蛋白,这些蛋白分别定位于鞭毛的近端或远端。这种旁系同源蛋白的分布模式和形成近端-远端非对称性的能力可能适用于大多数真核生物的鞭毛和纤毛。
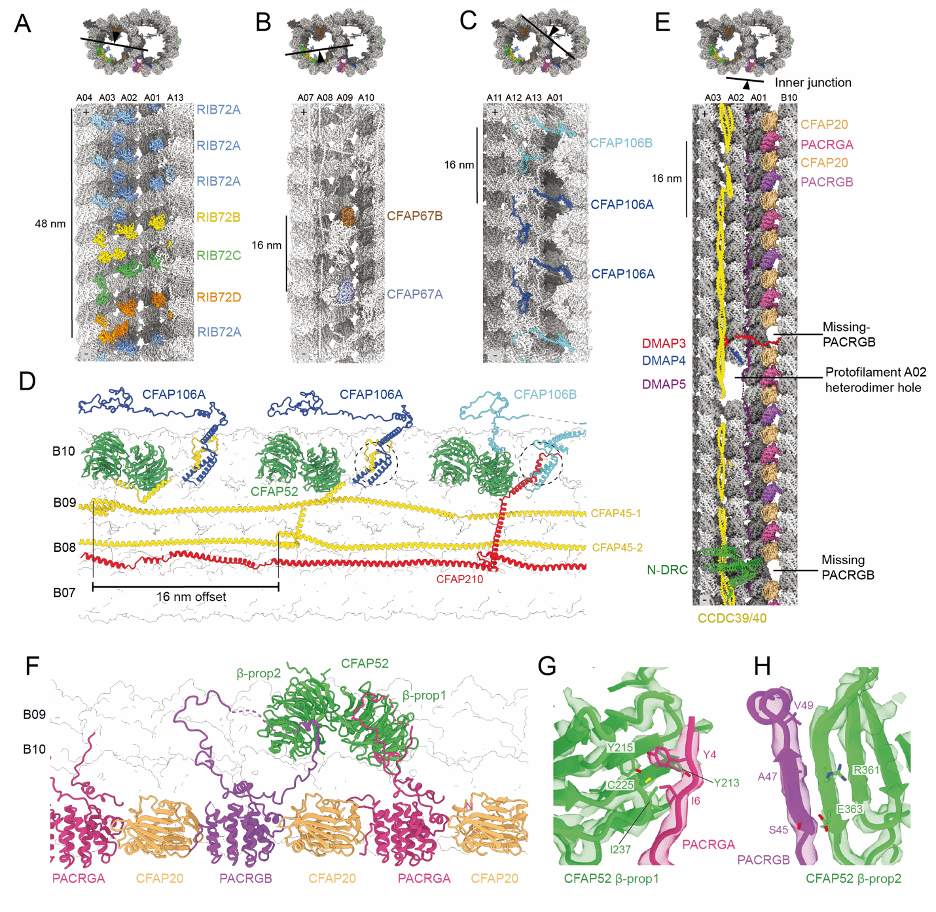
图 3:L. tarentolae 和 C. fasciculata DMT 的 旁系同源基因
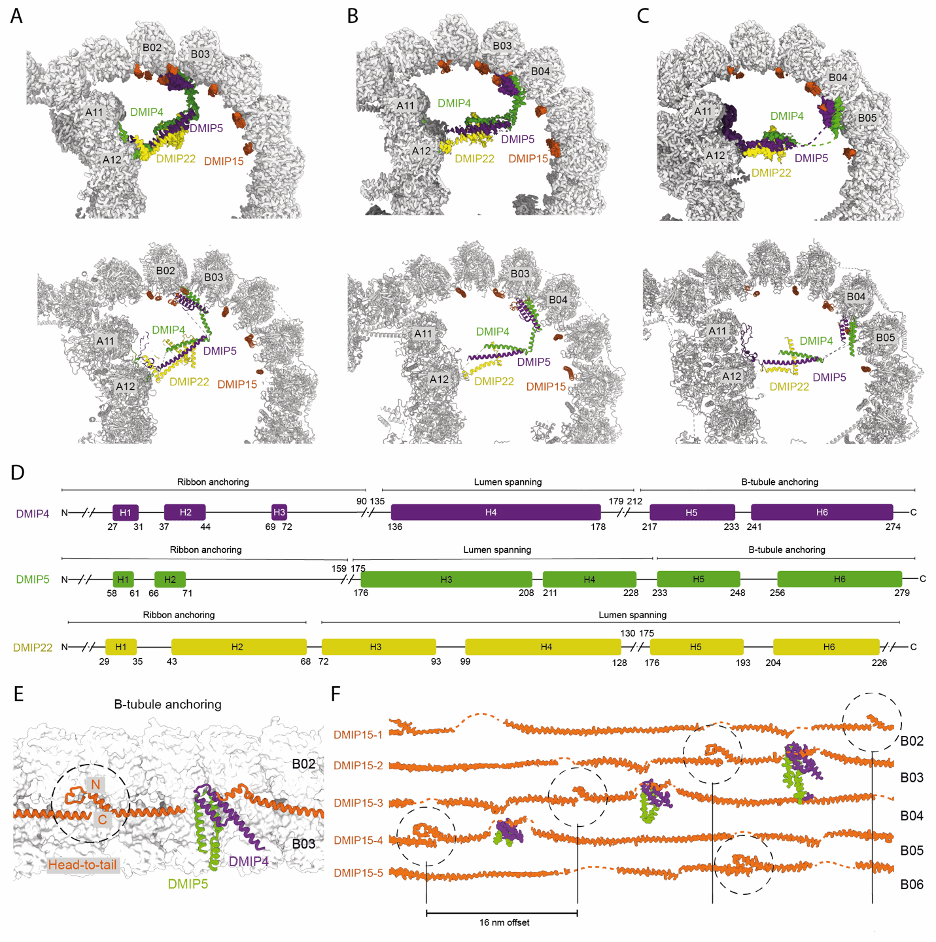
图 4:B-微管腔内的小桥(ponticulus)的结构基础
锥虫类 DMTs 的一个特征是 B-微管腔内存在的重复密度结构,这些结构被称为小桥(ponticuli),源自拉丁语。T. brucei DMTs 的 Cryo-ET 和 subtomogram averaging 显示在一个 48 nm 的重复单元内存在 3 个小桥【4】。这些小桥彼此相距 16 nm,并且都连接到 A-微管相同的原丝(A12)。与先前预期的每个小桥对应于不同的微管内蛋白(MIP)不同,我们发现每个小桥实际上都是同一个由 DMIP4、DMIP5 和 DMIP22 组成的三聚复合物形成【图 4】。它们共同形成一个细长的、主要由 α 螺旋构成的复合物。连接螺旋的柔性环提供了必要的可塑性,使其能够跨越 B-微管的多个结合位点。
我们进一步利用锥虫寄生虫的遗传可操作性(genetic tractability)以及它们的运动行为易于记录的特点,系统性地测试了每种已鉴定蛋白对寄生虫游动行为的影响。通过 CRISPR 基因编辑技术,我们在 Leishmania mexicana 中分别敲除了这些蛋白。随后,我们使用暗场视频显微镜(dark-field videomicroscopy)测量每个突变体的游动速度和方向性【图 5】。
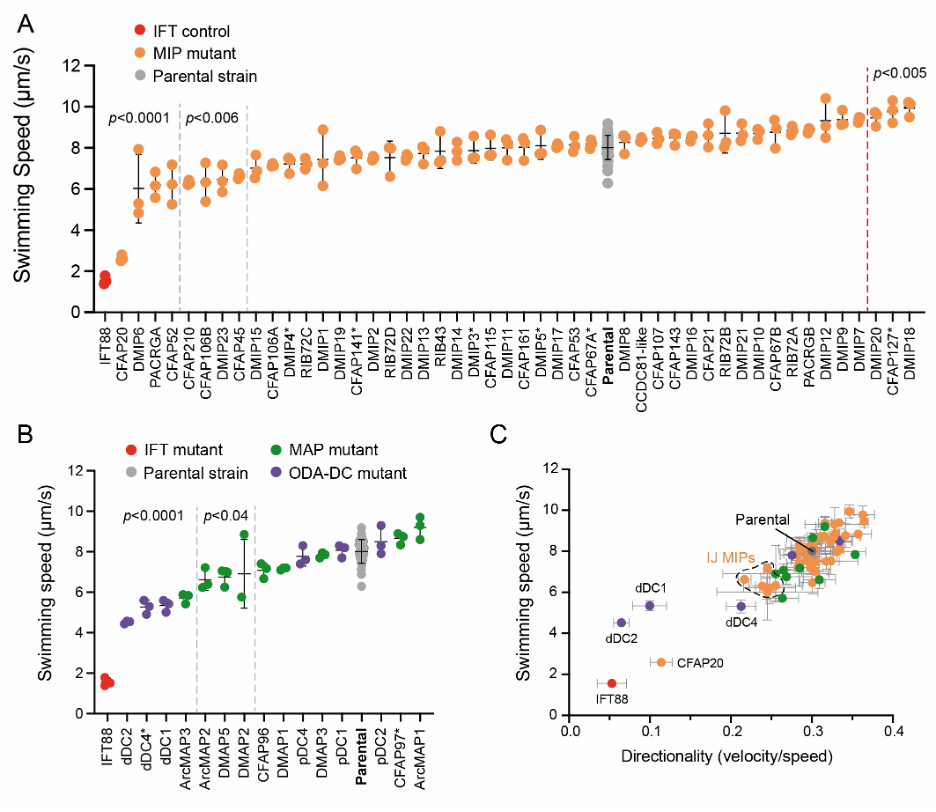
图 5:L. mexicana 敲除突变体的运动表型
数据分析显示,A-微管与 B-微管之间的内连接(inner junction)对基因突变的干扰高度敏感。大多数内连接 MIP 的基因缺失都会导致显著的游动速度下降。此外,我们通过基于结构的生物信息学分析,发现该区域包含一组在所有具有运动性鞭毛的生物中都保守存在的核心 MIPs,进一步强调了内连接区域对鞭毛运动的重要性。
综上所述,本研究的结构分析以分子水平解析了锥虫寄生虫双联微管的物种特异性特征,增强了我们对轴丝微管如何被 MIPs 调控和模式化的理解。通过系统性地评估每种结构验证蛋白对鞭毛运动的贡献,我们明确了内连接结构对于鞭毛正常运动的重要性。本研究采用了一种整合式方法,结合结构解析与系统性基因敲除分析,为揭示控制鞭毛摆动的保守机制以及锥虫寄生虫特异性机制提供了新的见解。
本文来源 [丁香学术]
推文原链接:https://mp.weixin.qq.com/s/OhawAwwnPv1mo-mXRi5d5g
福建农林大学分析测试中心
2026年1月30日







